" חלק ראשון
האנזים שמזרז את השלב הרביעי של מחזור הקרבס הוא α-keto glutarate dehydrogenase; אנזים זה הוא קומפלקס אנזים הדומה מאוד לפירובאט דהידרוגנאז. שניהם מורכבים מ- 48-60 חלבונים שבהם מוכרות שלוש פעילויות אנזימטיות שונות וגם בעלות אותם קופקטורים אנזימטיים; הם אנזימים דומים מאוד מכיוון שהם פועלים על מצעים דומים: הן פירובאט והן l "α-קטו גלוטרט, הם α- חומצות קטו. מנגנון הפעולה של שני הקומפלקסים האנזימטיים זהה.
ההתקפה של תיאמין פירופוספט על הקרבוניל (C = O) של "α-קטוגלוטראט, מוביל לדקרבוקסילציה שלו ונגזרת הנגזרת של קרבוקסיהידרוקסי פרופיל. עם ההעברה לאחר מכן לליפואמיד, מתרחש תהליך חיזור פנימי, שממנו מתקבל הנגזרות קרבוקסי ליפואמיד או ליפואמיד סוקסיניל.
לאחר מכן מגיב ה- succinam ליפומיד עם קואנזים A ונותן succinyl קו -אנזים A (שנמשך במחזור הקרבס) והליפואמיד המופחת אשר חמצן מחדש על ידי ה- FAD: ה- FADH2 שנוצר מתחמצן מחדש על ידי NAD + ומתקבל NADH. לפיכך, בשלב זה התרחשה חיסולו השני של פחמן מהשלד הפחמתי, בצורה של פחמן דו חמצני.
קבוצת אציל המקושרת לקואנזים A נמצאת בצורה מופעלת, כלומר יש לה תכולת אנרגיה גבוהה: לכן ניתן לנצל את האנרגיה של סוקסיניל קואנזים A.

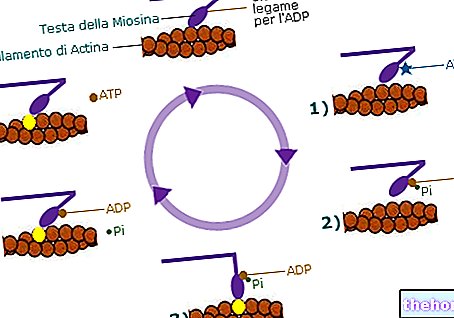
בשלב החמישי של מחזור הקרבס, סוכניל קואנזים A נתון לפעולה של succinyl thiokinase; הועלו שתי השערות לגבי אופן הפעולה שלו: נתאר רק אחת מהשתיים מכיוון שהיא המוסמכת ביותר. על פי השערה זו, סוציניל קואנזים A מותקף על ידי חנקן של היסטידין (Hys) של האנזים: קואנזים A משתחרר ומתווסף שמקורו בהיסטדין נוצר כתווך, כלומר האנזים סוקסיניל (או succinyl-Hys ); אורתופוספט פועל על ביניים זה, מה שמוביל לשחרור הסוקסינט ויצירת הפוספנזים. הפוספנזים, המותקף על ידי גונוזין דיפוספט (תוצר), מייצר גוואסנוזין טריפוספט (GTP) והאנזים משתחרר. מנקודת המבט האנרגטית GTP = ATP: הקשר המספק אנרגיה זהה בשני המינים (זהו הקשר האנהידריד בין פוספוריל Β לפוספוריל γ). במקרים מסוימים, GTP משמש כחומר בעל תכולת אנרגיה גבוהה אך בדרך כלל, GTP הופך ל- ATP על ידי פעולת האנזים נוקלאוזיד דיפוספו קינאז (NDPK); הוא אנזים הנמצא בתאים ומזרז את התגובה הבאה:
N1TP + N2DP → N1DP + N2TP
NiTP ® Nucleoside טריפוספט גנרי
NiDP ® נוקלאוזיד דיפוספט
זוהי תגובה הפיכה; במקרה שלנו זה קורה:
GTP + ADP → תוצר + ATP
לכן הוא יכול להמשיך ימינה או שמאלה אפילו לשינויים קטנים בריכוזי הריאגנטים.
אם מחזור הקרבס ממשיך במהירות כזו שתוביל לייצור ATP גבוה מהדרישה לאנרגיה, יש זמינות נדירה של ADP בעוד שיש הרבה ATP: התגובה המזרזת על ידי הדיפוספו קינאז נוקלאוזיד היא, אם כן, מופנה שמאלה (GTP מצטבר אם לקינאז הדיפוספו של נוקלאוזיד אין מצע מספיק כלומר ADP). GTP הוא אפוא אות לזמינות אנרגיה ולכן מאט את מחזור הקרבס.
השלב השישי של מחזור הקרבס מוביל להיווצרות הפומרט על ידי פעולתו של succinate dehydrogenase; אנזים זה נותן תגובה סטריאו -ספציפית כאשר הטרנס הבלתי רווי (הוא אלקן) נוצר תמיד, כלומר הפומרט (בעוד שאיזומר סיס הוא המליאט). Sucinate dehydrogenase נמצא על קרום המיטוכונדריה הפנימית, בעוד שכל האנזימים האחרים של מחזור הקרבס מפוזרים ברחבי המיטוכונדריה.
ל- succinate dehydrogenase יש FAD כקופקטור; הוא מעוכב על ידי אוקסלואצטט (עיכוב הפיד) בעוד שהוא מתפשט ופוומרט כמאפנן החיובי שלו (מפעיל) הפעיל שלו. בואו ננסה להבין מדוע, על ידי קפיצה לשלב האחרון של מחזור הקרבס. השלב האחרון של מחזור krebs דורש אנרגיה ולכן האפשרות היחידה להשיג אוקסלואצטט מהחולה היא שריכוז החולה גבוה מאוד: מאלט הוא אחד המטבוליטים עם הריכוז הגבוה ביותר בתאים. עובדה כי ריכוז האוקסלאצטט נשמר נמוך על ידי פעולת סינתז סיטראט. התגובה המזרזת על ידי succinate dehydrogenase היא, אם כן, תגובה להאכילה עצמית וזו הדרך היחידה לגרום להפיכת המלט לאוקסולאצטט להתרחש.
ריכוז המלט המיטוכונדריאלי חייב להיות תואם את ריכוז המלט הציטופלסמי: רק כאשר ריכוז המלט המיטוכונדריאלי גבוה עד כדי להבטיח את הפיכת המלט לאוקסולאצטט (במחזור הקרבס), ניתן להשתמש במלט גם ב- דרכים אחרות (שהן ציטופלזמה): בציטופלזמה ניתן להפוך את המלט לאוקסולאצטט שממנו ניתן להשיג אספרטט באמצעות פעולת GOT (זהו טרנסמינאז) או גלוקוז באמצעות גלוקונאוגנזה.
אנו חוזרים לשלב השביעי של מחזור הקרבס הוא מזורז על ידי האנזים fumarasi: מים מתווספים בצורה סטריאו-ספציפית ליצירת L-malate.
בשלב האחרון של מחזור קרבס, עליו כבר דיברנו, הפעולה של malate dehydrogenase. אנזים זה משתמש במולקולת NAD + לפעולתו הקטליטי.
סיכמנו אפוא את תיאור השלבים השונים של מחזור הקרבס.
מחזור הקרבס הפיך לחלוטין.
כדי להגדיל את מהירות מחזור הקרבס, ניתן להגדיל את ריכוז המטבוליטים הקיימים באותו מחזור; אחת האסטרטגיות להגדלת מהירות מחזור הקרבס מורכבת מהפיכת חלק מהפירובט הנכנס למיטוכונדריה לאוקסולאצטט (על ידי פעולת פירובט קרבוקסילאז) ולא הופך את כל זה לאצטיל קואנזים A: ובכך מגביר את ריכוז האקסלואצטט אשר הוא מטבוליט של מחזור הקרבס ולכן מגביר את מהירות המחזור כולו.
במחזור הקרבס שלושה NAD + מומרים לשלושה NADH ואחד FAD ל- FADH2, יתר על כן מתקבל GTP: על ידי תקשור הכוח המפחית המתקבל ממחזור הקרבס, ייוצר ATP נוסף; בשרשרת הנשימה, העוצמה המפחיתה מועברת מ- NADH ו- FADH2 לחמצן: העברה זו נובעת מסדרת אנזימים הממוקמים על הממברנה המיטוכונדריאלית אשר בפעולתם מביאים לייצור ATP.
התהליכים של שרשרת הנשימה הם תהליכים אקסרגוניים והאנרגיה המשוחררת משמשת לייצור ATP; מטרת התא היא לנצל את התהליכים האקסרגוניים כדי לגרום לסינתזה של ATP להתרחש. עבור כל מולקולת NADH שנכנסת לשרשרת הנשימה מתקבלים 2.5 מולקולות ATP ועל כל FADH2 מתקבלות 1.5 מולקולות ATP; גיוון זה נובע מכך ש- FADH2 נכנס לשרשרת הנשימה ברמה נמוכה יותר מ- NADH.
עם הכוח המפחית של חילוף החומרים האירובי, 30-32 ATP (219-233 קק"ל / מול) מתקבלים ביעילות של כ -33% (יעילות המטבוליזם האנאירובי היא כ -2%).