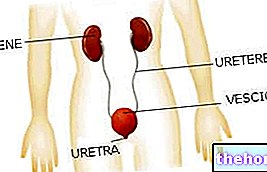
כדי שתהיה התאמה בין המידע של הפולינוקלאוטיד לזה של הפוליפפטיד, יש קוד: הקוד הגנטי.
ניתן להציג את המאפיינים הכלליים של הקוד הגנטי כדלקמן:
הקוד הגנטי בנוי משלישות, ונטול סימני פיסוק פנימיים (Crick & Brenner,).
הוא "פוענח באמצעות" מערכות תרגום תאים פתוחים "(Nirenberg & Matthaei, 1961; Nirenberg & Leder, 1964; Korana, 1964).
זה מאוד מנוון (מילים נרדפות).
ארגון טבלת הקוד אינו מקרי.
שלישיות "שטויות".
הקוד הגנטי הוא "סטנדרטי", אך לא "אוניברסלי".
בהסתכלות על טבלת הקוד הגנטי, יש לזכור שהיא מתייחסת לתרגום של "RNAm לפוליפפטיד, שעבורו בסיסי הנוקלאוטיד המעורבים הם A, U, G, C. הביוסינתזה של שרשרת פוליפפטיד היא התרגום של רצף הנוקלאוטיד ברצף חומצת אמינו.

לכל שלישיית בסיס של ה- RNAm, הנקראת קודון, יש את הבסיס הראשון בעמודה השמאלית, השני בשורה העליונה, השלישי בעמודה הימנית. ניקח לדוגמא טריפטופן (כלומר נסה) ואנו רואים שהקודון המתאים להיות, לפי הסדר, UGG. למעשה, הבסיס הראשון, U, כולל את כל שורת הקופסאות בחלקו העליון; בכך G מזהה את התיבה הימנית ביותר ואת השורה הרביעית של התיבה עצמה, שם אנו מוצאים את כתוב נסה. באופן דומה, כדי לסנתז את הטטראפפטיד Leucine-Alanine-Arginine-Serína (סמלים Leu-Ala-Arg-Ser) אנו יכולים למצוא את הקודונים UUA-AUC-AGA-UCA בקוד.

עם זאת, בנקודה זו יש לציין כי כל חומצות האמינו של הטטרפפטיד שלנו מקודדות (בניגוד לטריפטופן) על ידי יותר מקודון אחד. לא במקרה בדוגמה שדיווחנו זה עתה בחרנו בקודונים המצוינים. יכולנו לקודד את אותו טריפפטיד ברצף RNAm שונה, כגון CUC-GCC-CGG-UCC.
בתחילה, העובדה שחומצת אמינו אחת התאימה ליותר משלישייה קיבלה משמעות של אקראיות, המתבטאת גם בבחירת מונח הניוון של הקוד, המשמשת להגדרת תופעת המילה הנרדפת. מאידך גיסא, נתונים מסוימים מצביעים על כך שזמינותם של מילים נרדפות המתייחסות ליציבות שונה של המידע הגנטי איננה מקרית כלל. נראה כי הדבר מאושר גם על ידי מציאת ערך שונה של יחס A + T / G + C בשלבי האבולוציה השונים. לדוגמה, בפרוקריוטים, שבהם הצורך בשונות אינו מסופק על ידי כללי המנדליזם והניאו-מנדליזם, יחס A + T / G + C נוטה לעלות. היציבות הנמוכה כתוצאה מכך, מול מוטציות, מספקת יותר הזדמנויות לשונות אקראיות ממוטציה גנטית.
באיקריוטים, בפרט בתאים רב תאיים, בהם יש צורך שהתאים של האורגניזם היחיד ישמרו על אותה ירושה תורשתית, יחס A + T / G + C ב- DNA נוטה לרדת, מה שמקטין את ההזדמנות למוטציות גנים סומטיים. .
קיומם של קודונים נרדפים בקוד הגנטי מעורר את הבעיה, כאמור, של ריבוי או לא של anticodons ב- RNAt.
זה בטוח שיש לפחות RNAt אחד לכל חומצת אמינו, אך לא בטוח אם RNAt אחד יכול להיקשר לקודון בודד, או שהוא יכול לזהות מילים נרדפות באדישות (במיוחד כאשר אלה שונות רק מהבסיס השלישי).
אנו יכולים להסיק כי יש בממוצע שלושה קודונים נרדפים לכל חומצת אמינו, בעוד שאנטי -קודונים הם לפחות אחד, ולא יותר משלושה.
נזכיר כי גנים מיועדים למתיחות בודדות של רצפי פולינונוקלאוטיד ארוכים מאוד של DNA, ברור כי בהכרח יש להכיל את ההתחלה והסוף של הגן היחיד בזיכרון.
ביו -סינתזה של חלבונים
בחלקים שונים של ה- DNA יש פתיחת השרשרת הכפולה והסינתזה של סוגי ה- RNA השונים.
במהלך שלב הטעינה, ה- RNAt נקשר לחומצות האמינו (שהופעלו בעבר על ידי ה- ATP ועל ידי האנזים הספציפי). ה"מכונה "הביו -סינתטית אינה מסוגלת" לתקן "tRNA טעונים בצורה לא נכונה.
לאחר מכן ה- RNAr מתפצל לשתי יחידות המשנה, ועל ידי קישור לחלבונים הריבוזומליים, נוצר הרכבה של הריבוזומים.
ה- RNAm, העובר דרך הציטופלזמה, נקשר לריבוזומים, יוצר את הפוליסום. כל ריבוזום, הזורם על השליח, מארח בהדרגה את ה- RNAt המשלים את הקודונים היחסיים, לוקח את חומצות האמינו ומחבר אותן לשרשרת הפוליפפטיד בהתהוות.
ה- RNAt היציב יחסית נכנס שוב למחזור הדם. הריבוזומים משמשים גם הם שוב, ומשחררים את הפוליפפטיד שכבר הורכב.
השליח, פחות יציב מכיוון שכולו חד -צמדי, נחלק (על ידי הריבונוקלאז) לתוך הריבונוקליאוטידים המרכיבים.
המחזור ממשיך אפוא, ומסנתז בזה אחר זה את הפוליפפטידים על ה- RNAs שליחים המסופקים על ידי התעתיק.