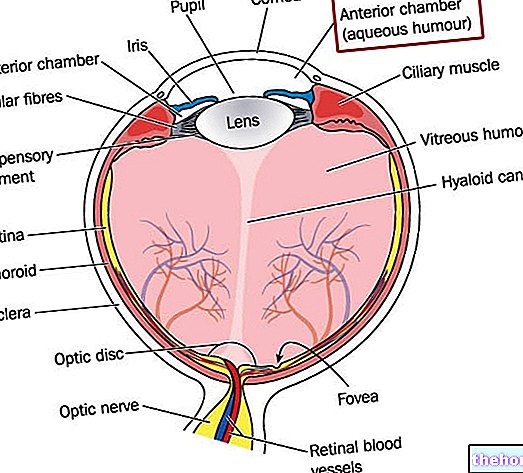
חשיבותו של מיוזה
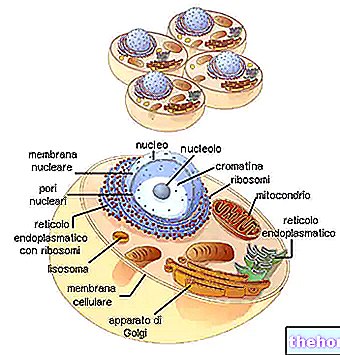
בהקשר של אורגניזם רב -תאי יש צורך שלכל התאים (כדי לא לזהות זה את זה כזר) יש את אותה ירושה תורשתית. הדבר מושג על ידי מיטוזה, חלוקת כרומוזומים בין תאי הבת, שבהם "שוויון הגנטי המידע מובטח על ידי מנגנון צמצום ה- DNA, בהמשכיות התא שעוברת מהזיגוטה לתאים האחרונים של האורגניזם, במה שמכונה הקו הסומטי של דורות התא.
עם זאת, אם אותו מנגנון היה מאומץ בדור הצאצאים, המין כולו נוטה להיות מורכב מאנשים שווים גנטית. היעדר השתנות גנטית כזו עלולה לפגוע בקלות בשרידות המין עם שינוי תנאי הסביבה. לכן יש צורך בכך שהמין, בהקשר של שונות החומר הגנטי שהוא מודה בו, עשוי לעורר מבחר מחדש, ערבוב, לא בהקשר של האורגניזם היחיד, אלא במעבר מדור לדור. הדבר נעשה על ידי תופעות המיניות ומנגנון חלוקת התא המסוים הנקרא מיוזה.

מהי מיוזה
מיוזה מתרחשת רק בתאים של חיידקים. כאשר סדרה ארוכה של חלוקות מיטוטיות הכפילה מספיק את מספר תאי הנבט הזמינים, האחרונים נכנסים למיוזה, ובכך מכינים את הגמטות. גמטות, המתמזגות להפריה, מאגדות את החומר הכרומוזומלי שלהן. אם הגמטאים היו דיפלואידים, בדומה לתאים האחרים של האורגניזם, ההתמזגות שלהם בזיגוטה הייתה נותנת לילדים בעלי מורשת 4n; אלה היו נותנים 8n ילדים וכן הלאה.
כדי לשמור על מספר הכרומוזומים של המין קבוע, על הגמטות להיות הפלואידיות, כלומר עם מספר n במקום 2n של כרומוזומים. זה מושג עם מיוזה.
ניתן להבין את מיוזיס כהמשכה של שתי חטיבות מיטוטיות מבלי שהשתלבות צמצום.
בכל אחת משתי החטיבות הרצופות, שמקורן בארבעה תאים הפלואידים מתא נבט דיפלואידי, יש רצף של פרופאז, מטאפאזה, אנאפאזה, טלופאז וציטודיאז.
עם זאת, הנבואה של החלוקה המיוטית הראשונה מסובכת במיוחד, ומביאה לרצף רגעים שלוקחים את השמות המתאימים של לפפטן, זיגוטן, פאצ'יטן, דיפלוטן ודיאקינזה.
אנו מתייחסים לרגעים אלה אחד אחד, בעקבות התנהגות זוג כרומוזומים בודד.
לפטין. זוהי תחילתה של מיוזה. הכרומוזומים מתחילים לראות זה את זה, עדיין אינם ספיראליים במיוחד.
זיגוטן. הכרומוזומים מזוהים בצורה ברורה יותר, ונדמה כי הכרומוזומים ההומולוגיים מתקרבים. (נזכיר כי החוטים הנוטים להתקרב, מקבילים זה לזה, הם 4: שני כרומטידים לכל אחד משני הכרומוזומים ההומולוגיים).
פצ'יטן. ארבעת החוטים הכרומטידים נדבקים לכל אורכו, מחליפים משיכות, על ידי שבירה וריתוך.
דיפלוטן. ככל שהספירלה ולכן ההתעבות גוברת, הכרומוזומים נוטים להניח את האינדיבידואליות הנפרדת שלהם: כאשר כל צנטרומר מתחבר לגדיל כפול.
הנקודות בהן התרחשה ההחלפה על ידי שבירה וריתוך (chiasma) עדיין מחזיקות את החוטים (כרומונמות) יחד בחלקים שונים. ארבעת הכרומנות, המחוברות בזוגות על ידי הצנטרומרים והדבקות שונות בשיאסימות, יוצרות את הטטרודות.
דיאצינזה. הטטרדות נוטות לסדר את עצמן בקו המשווה של הציר; הממברנה הגרעינית נעלמה; ההפרדה של הצנטרומרים מתחילה. כשזה קורה, הכרומוזומים, שכבר מאוחדים בצ'יזמים, נפרדים.
לאחר המטאפזה הבאה, שני הצנטרומרים (שטרם הוכפלו) נודדים לכיוון הקטבים הנגדים של הציר.
לאחר מכן, ברצף מהיר, האנאפאזה, הטלופאז והציטודיאזיס של החטיבה הראשונה, ומיד לאחר מכן החלוקה השנייה.
בעוד שאחרי המטאפאזה של החלוקה הראשונה הצנטרומים נדדו אל קטבי הציר וגררו שני חוטים, במטאפזה השנייה כל צנטרומר מוכפל. שני התאים הנובעים מהחלוקה הראשונה קיבלו n צנטרומרים עם 2n נימים, אך חלוקתם לאחר מכן מביאה ל -4 תאים, לכל אחד נימים (כלומר, בשלב זה, n כרומוזומים).
תכנית כללית זו מסבירה שלוש תופעות שונות ומקבילות:
- צמצום הערכה הכרומוזומלית מהדיפלואיד (2n) של ה"אורגניזם ל "הפלואיד (n) של הגמט.
- הייחוס האקראי לגאמט של הכרומוזום אחד או אחר, ממוצא אימהי או אבהי.
- חילופי חומרים גנטיים בין כרומוזומים הומולוגיים ממוצא אבהי ואימהי (עם ערבוב של החומר הגנטי, לא רק ברמה של כרומוזומים שלמים, אלא גם בתוך הכרומוזומים עצמם).