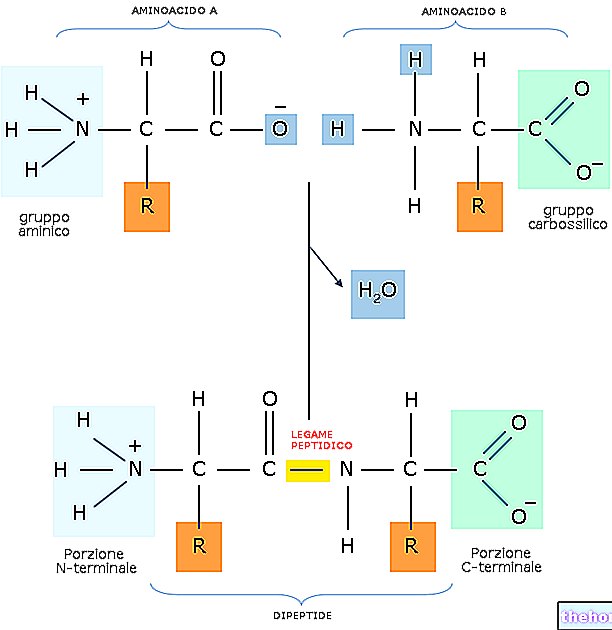
כדי לדבר על עשרים חומצות האמינו המרכיבות את מבני החלבון והשינויים, יהיה צורך לתאר לפחות שתים עשרה מסלולים מטבוליים מיוחדים.
אבל מדוע התאים משתמשים בכל כך הרבה מסלולים מטבוליים הדורשים אנרגיה (למשל כדי לשחזר את האתרים הקטליטיים של אנזימים), כל אחד עם ירושה אנזימטית, כדי להטיל חומצות אמינו? כמעט מכל חומצות האמינו ניתן להשיג באמצעות מסלולים מיוחדים מטבוליטים המשמשים בחלקם ליצירת אנרגיה (למשל באמצעות גלוקונאוגנזה ומסלול של גופי קטון) אך מעל לכל מובילים להיווצרות קומפלקס מורכב. מולקולות, בעלות מספר גבוה של אטומי פחמן (למשל מפנילאלנין וטירוזין, נוצרים הורמונים בבלוטת האדרנל המתמחים למטרה זו); אם מצד אחד יהיה פשוט לייצר אנרגיה מחומצות אמינו, מצד שני יהיה מסובך לבנות מולקולות מורכבות החל ממולקולות קטנות: קטבוליזם של חומצות אמינו מאפשר להם לנצל את השלד שלהם כדי להשיג מינים גדולים יותר.
שניים או שלושה ק"ג של חומצות אמינו מתפרקות מדי יום על ידי אדם בריא: 60-100 גרם מהן נובעות מהחלבונים שהוכנסו לתזונה אך יותר משני ק"ג מתקבלים מהמחזור הרגיל של החלבונים המהווים חלק בלתי נפרד מהאורגניזם ( חומצות אמינו של חלבונים אלה, אשר ניזוקו מתהליכי חיזור, מוחלפות באחרים ומתמוטטות).
חומצות אמינו נותנות תרומה אנרגטית במונחים של ATP: לאחר הסרת קבוצת α-אמינו, השלד הפחמני הנותר של חומצות אמינו, בעקבות טרנספורמציות מתאימות, יכול להיכנס למחזור הקרבס. יתר על כן, כאשר אספקה של חומרים מזינים חסרה וכמות הגלוקוז יורדת, גלוקונאוגנזה מופעלת: אומרים כי חומצות אמינו גלוקונאוגנטיות הן אלה שאחרי שינויים מתאימים ניתן להכניס אותן לגלוקונאוגנזה; חומצות אמינו גלוקונאוגניות הן אלה שניתן להפוך אותן פירובאט או בפומאראט (ניתן להפוך פומרט למלט שעוזב את המיטוכונדריון ובציטופלזמה הופך לאוקסולאצטט שממנו ניתן להשיג פוספנול פירובאט). חומץ-אצטט.
זה שתואר זה היבט חשוב מאוד מכיוון שחומצות אמינו יכולות לתקן מחסור בסוכר במקרה של צום מיידי; אם הצום נמשך, לאחר יומיים חילוף החומרים של השומנים מתערב (מכיוון שלא ניתן לתקוף את מבני החלבון יותר מדי), בשלב זה מכיוון שגלוקונאוגנזה מוגבלת מאוד, חומצות השומן הופכות לאצטיל קואנזים A וגופי קטון. מצום נוסף, המוח מסתגל גם לשימוש בגופי הקטון.
העברת קבוצת α-אמינו מחומצות אמינו מתרחשת באמצעות תגובת טרנסמינציה; האנזימים שמזרזים את התגובה הזו, הם אומרים, למעשה, טרנסמינאזות (או אמינו טרנספראז). אנזימים אלה משתמשים בקופקטור אנזימטי בשם pyridoxal phosphate, המתערב בקבוצת האלדהיד שלו. פוספט פירידוקסאל הוא תוצר הפוספורילציה של פירידוקסין שהוא ויטמין (B6) המצוי בעיקר בירקות.
לטרנסמינאזות יש את המאפיינים הבאים:
ספציפיות גבוהה לזוג α קטוגלוטראט-גלוטמט;
הם נקראים על שם הזוג השני.
אנזימי טרנסמינאז כוללים תמיד את צמד α ketoglutarate-glutamate והם נבדלים בהתאם לזוג השני המעורב.
דוגמאות:
L "טרנסמינאז אספרטט כלומר GOT (Glutamate-Ossal acetate Transaminase): האנזים מעביר את קבוצת α-אמינו מאספרטט ל- α-ketoglutarate, תוך קבלת אוקסלואצטט וגלוטמט.
L "אלנין טרנסמינאז כלומר GTP (Glutamate-Pyruvate Transaminase): האנזים מעביר את קבוצת α-אמינו מ"אלנין ל- "α-ketoglutarate, תוך קבלת פירובאט וגלוטמט.
הטרנסמינאזות השונות משתמשות ב- α-ketoglurate כמקבץ של קבוצת האמינו של חומצות האמינו והופכות אותו לגלוטמט; בעוד שחומצות האמינו הנוצרות משמשות את מסלול גופי הקטון.
תגובה מסוג זה יכולה לקרות לשני הכיוונים מכיוון שהם שוברים ויוצרים קשרים עם אותו תכולת אנרגיה.
הטרנסמינאזות נמצאות הן בציטופלזמה והן במיטוכונדריון (הן פעילות בעיקר בציטופלזמה) והן שונות בנקודה האיזואלקטרית שלהן.
הטרנסמינאזות מסוגלות גם decarboxylate חומצות אמינו.
חייבת להיות דרך להפוך את הגלוטמט בחזרה ל- α-ketoglutarate: הדבר נעשה על ידי דימינציה.
שם גלוטמט דהידרוגנאז זהו אנזים המסוגל להפוך גלוטמט ל- α-ketoglutarate, ולכן, להפוך את קבוצות האמינו של חומצות האמינו הנמצאות בצורת גלוטמט לאמוניה. מה שמתרחש הוא תהליך חמצון שעובר דרך האמצע הבינוני α-amino: אמוניה ו- α-ketoglutarate משתחררים וחוזרים למחזור הדם.
לאחר מכן, סילוק קבוצות האמינו של חומצות האמינו עובר דרך הטרנסאמינאזות (השונות בהתאם למצע) וגלוטמט הדהידרוגנאז, הקובע את היווצרות האמוניה.
ישנם שני סוגים של גלוטמט דיהידרוגנאז: ציטופלסמה ומיטוכונדריאלית; הקופקטור, שהוא גם קוסובסטרט של האנזים הזה הוא NAD (P) +: גלוטמט דהידרוגנאז משתמש ב- NAD + או NADP + כמקבל הפחתת כוח. הצורה הציטופלסמית מעדיפה, אם כי לא באופן בלעדי, NADP + ואילו הצורה המיטוכונדרית מעדיפה NAD +. הצורה המיטוכונדרית נועדה להיפטר מקבוצות אמינו: היא מובילה להיווצרות אמוניה (המהווה מצע לאנזים מיוחד במיטוכונדריון) ו- NADH (הנשלח לשרשרת הנשימה). הצורה הציטופלסמית פועלת בכיוון ההפוך כלומר, היא משתמשת באמוניה ו- α-ketoglutarate כדי לתת גלוטמט (שיש לו יעד ביוסינתטי): תגובה זו היא ביוסינתזה רדוקטיבית והקופקטור המשמש הוא NADPH.
גלוטמט דהידרוגנאז פועל כאשר יש צורך להיפטר מקבוצות האמינו של חומצות אמינו כגון אמוניה (דרך שתן) או כאשר יש צורך בשלדי חומצות אמינו לייצור אנרגיה: לאנזים זה יהיו אפוא כמאפננים שליליים את המערכות המהוות אינדיקציה. בעל זמינות טובה של אנרגיה (ATP, GTP ו- NAD (P) H) וכמאפננים חיוביים, המערכות המצביעות על צורך באנרגיה (AMP, ADP, GDP, NAD (P) +, חומצות אמינו והורמוני בלוטת התריס).
חומצות אמינו (בעיקר לאוצין) הן מאפניות חיוביות של גלוטמט דיהידרוגנאז: אם חומצות אמינו נמצאות בציטופלזמה, ניתן להשתמש בהן לסינתזת חלבונים, או להיפטר מהן מכיוון שלא ניתן לצבור אותן (זה מסביר מדוע חומצות אמינו הן מאפניות חיוביות. ).
סילוק אמוניה: מחזור אוריאה
דגים נפטרים מאמוניה על ידי החדרתם למים דרך הזימים; ציפורים הופכות אותו לחומצת שתן (שהיא תוצר עיבוי) ומחסלות אותו בצואה. בואו נראה מה קורה בבני אדם: אמרנו שגלוטמט דהידרוגנאז ממיר גלוטמט ב- α- ketoglutarate ואמוניה אך לא אמרנו שזה קורה רק במיטוכונדריה של הכבד.
תפקיד בסיסי של סילוק אמוניה, באמצעות מחזור האוריאה, ממלא טרנסמינאז מיטוכונדריאלי.

פחמן דו חמצני, בצורת יון ביקרבונט (HCO3-), מופעל על ידי קופקטור הביוטין היוצר ביוטין קרבוקסי המגיב עם אמוניה לתת חומצה קרבמית; התגובה הבאה משתמשת ב- ATP כדי להעביר פוספט לחומצה הקרבמית היוצרת פוספט קרביל ו ADP (ההמרה של ATP ל- ADP היא הכוח המניע להשגת קרבוקסיביוטין). שלב זה מזורז על ידי carbamyl phosphate synthetase ומופיע במיטוכונדריה. פוספט קרבמיל ואורניטין הם מצעים לאנזים אורניטין טרנס קרבמילאז מה שהופך אותם לסיטרולליין; תגובה זו מתרחשת במיטוכונדריה (הפטוציטים). הציטרולליין המיוצר עוזב את המיטוכונדריון ובציטופלזמה הוא תחת "פעולה של"ארגינין סוקסינט סינתטאז: יש מיזוג בין השלד הפחמני של ציטרוללין לזה של אספרטט באמצעות התקפה נוקלאופילית וסילוק מים לאחר מכן. האנזים סינתטאז ארגינין סוקסינט דורש מולקולת ATP כך שיש צימוד אנרגטי: הידרוליזה של ה- ATP ל- AMP ופירופוספט (האחרון מומר לאחר מכן לשתי מולקולות של אורתופוספט) מתרחשת על ידי גירוש מולקולה d "מים מהמצע. ולא על ידי פעולת מי המדיום.
"האנזים הבא הוא ה"ארגנין סוקסינאז: אנזים זה מסוגל לפצל את ארגינין סוקסינט לארגנין ולפומרט בתוך הציטופלזמה.
מחזור האוריאה מושלם על ידי האנזים arginase: מתקבלים אוריאה ואורניטין; אוריאה נפטרת על ידי הכליות (שתן) ואילו אורניטין חוזר למיטוכונדריה ומחדש את המחזור.
מחזור האוריאה נתון לאפנון עקיף על ידי ארגינין: הצטברות הארגנין מצביעה על כך שיש להאיץ את מחזור האוריאה; אפנון הארגנין הוא עקיף מכיוון שהארגינין מווסת באופן חיובי את האנזים סינתטאז אצטיל גלוטמט. האחרון מסוגל להעביר קבוצת אצטיל על החנקן של גלוטמט: נוצר N- אצטיל גלוטמט המהווה אפנן ישיר של האנזים סינתטאז carbamyl-phospho.
ארגינין מצטבר כמטבוליט של מחזור האוריאה אם ייצור פוספט קרבמיל אינו מספיק כדי להיפטר מהאורניטין.
אוריאה מיוצרת רק בכבד אך ישנם אתרים נוספים בהם מתרחשות התגובות הראשוניות.
המוח והשרירים משתמשים באסטרטגיות מיוחדות לחיסול קבוצות אמינו. המוח משתמש בשיטה יעילה מאוד בה משתמשים באנזים גלוטמין סינתטאז ואנזים גלוטמאז: הראשון קיים בנוירונים, ואילו השני נמצא בכבד. מנגנון זה יעיל מאוד משתי סיבות:
שתי קבוצות אמינו מועברות מהמוח אל הכבד ברכב אחד;
גלוטמין הרבה פחות רעיל מאשר גלוטמט (גלוטמט מבצע גם העברה עצבית ואסור לחרוג מהריכוז הפיזיולוגי).
אצל דגים מנגנון דומה מביא את קבוצת האמינו של חומצות האמינו לזימים.
מהשריר (השלד והלב), קבוצות האמינו מגיעות לכבד דרך מחזור הגלוקוז-אלאנין; האנזים המעורב הוא טרנסמינאז גלוטמין-פירובאט: הוא מאפשר טרנספוזיציה של קבוצות אמינו (בצורת גלוטמט), הפיכת פירובאט לאלאנין, ובמקביל גלוטמט ל- α-ketoglutarate בשריר, ומזרז את תהליך הפוך בכבד.
לטרנסמינאזות עם משימות או עמדות שונות יש גם הבדלים מבניים וניתנים לקביעה על ידי אלקטרופורזה (יש להם נקודות איזואלקטריות שונות).
נוכחותם של טרנסמינאזות בדם יכולה להיות סימפטום של פגיעה בכבד או בלב (כלומר פגיעה ברקמות בכבד או בתאי הלב); הטרנסמינאזות נמצאות בריכוזים גבוהים מאוד הן בכבד והן בלב: באמצעות אלקטרופורזה ניתן לקבוע האם הנזק אירע בכבד או בתאי הלב.



.jpg)