איגוד (או "חיבור" או "קישור")
עד כה טיפלנו במעבר הדיהיבריד (או הפוליהיבריד) המנדלי מתוך הנחה שזוגות האלל השונים נמצאים למעשה על זוגות כרומוזומים הומולוגיים שונים. להגיע למאה כרומוזומים), בעוד שמספר הגנים ניתן לספור בעשרות אלפים.
העובדה שהדמויות שבחר מנדל לניסוייו נפרדו באופן עצמאי (מבלי לבלבל לכן את החישובים בהתפלגות הפנוטיפים ב- F2 של הצלב הפוליהיברידי) היה סיכוי בר מזל. אם נמצאו שני זוגות אללים על לוקוסים סמוכים, החוק ש היה עוקב ייקרא חוק ההתאגדות.
בידיעה שלדמויות רבות מאוד יש את המיקום שלהן על כרומוזום אחד וכי הן הזוגות הכרומוזומליים המפרידים באופן עצמאי במיוזה, ניתן להבין היטב באיזו תדירות קורה ששני זוגות של תווים, אם היו קשורים לכרומוזום אצל ההורה. אורגניזם, להישאר קשור. באותה מידה גם בגמטא ולכן באורגניזם שאליו יביא חומר גנטי משלו.
כך אנו רואים ש"האגודה מייצגת "חריגה", רחוקה מלהיות נדירה, לעצמאות המובנית בחוק השלישי של מנדל.
החלפה או "מעבר חוצה" ושילוב
אם כבר מדברים על מיוזה ציינו שיש שני רגעים שונים של ערבוב של החומר הגנטי: האחד הוא של הפרדת הכרומוזומים בגמטים, וזהו זה שנצפה על ידי מנדל.
הרגע השני, שלמעשה הוא הקודם, הוא זה שבו ארבעת הכרומטידים של כל זוג כרומוזומים הומולוגיים מחליפים הדדית תכונות זהות. שהחלפה מתבצעת היא פרופורציונלית, לקירוב ראשון, לאורכו של הכרומוזום, ובכרומוזומים ארוכים יותר יכולה להיות אפילו יותר מההחלפה.
ניתן לזהות את התופעה באופן ציטולוגי, על ידי התבוננות במספר מספיק של מיאוות מתחת למיקרוסקופ.
שיעור ההתאמה מחדש הוא הקצב שבו כל שתי דמויות שהיו קשורות בדור ההורים מתאחדות באופן שונה ב- F2.
אם שני הלוקוסים רציפים לחלוטין, ההסתברות כי שיאזם יפריד ביניהם תהיה אפסית כמעט. שיעור רקומבינציה יהיה: n רקומביננטיות. אם שני לוקוסים נמצאים על שני כרומוזומים שונים, קצב הרקומבינציה יהיה 0.5 (הסתברות שווה, עבור שתי תווים שחוברו בדור ה- P, כדי למצוא את עצמם באופן אקראי ב- F2). קצב השילוב מחדש יכול להשתנות בין 0.0 ל -0.5. למרחקים קטנים בכרומוזום, המרחק וקצב השילוב הינם ביחס ישר. למרחקים ארוכים יותר קיימת האפשרות כי מתקיימים שני חילופי דברים בין שני לוקוסים. כעת ייראה ברור ששני גורמים המופרדים על ידי שני חילופים קשורים שוב. ברור, בנקודה זו, שהפרופורציות בין מרחק הלוקוסים לבין ההסתברות לשילוב מחדש הולכת לאיבוד.
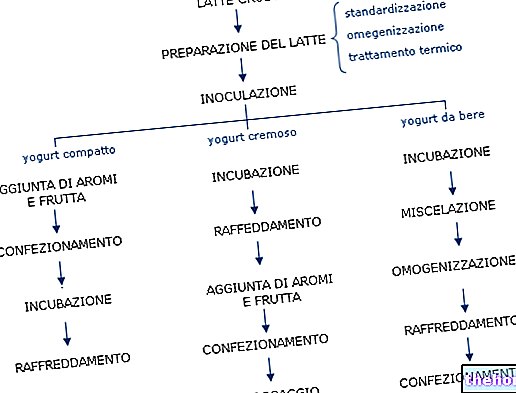
קשרים שנמצאו קשורים לאותו כרומוזום מהווים 'קבוצות איגוד'. לוקוסים רחוקים מאוד עשויים להיות בעלי הסתברות כזו של הפרדה באמצעות חילופי שהם מתנהגים כעצמאים, אך כל אחד מהם יקושר, עם שיעור רקומבינציה נמוך יותר, לאוקי הביניים.
כאשר שיעורי החיבור מחדש בין זוגות גנים רבים בתוך קבוצת התאחדות ידועים, ניתן להתחיל בבניית 'מפות גנטיות'. זכור כי המרחק בין שני גנים (a ו- b) מתבטא בקצב הרקומבינציה וכי המרחק של a מהגן השלישי c יכול להיות הסכום או ההבדל ביחס למרחקו מ- b, זה אפשרי לשחזר מפה של המרחקים ההדדיים, שתהיה המפה הגנטית בתוך קבוצת ההתאחדות, כלומר של אותו כרומוזום.
כעת עלינו לשקול באופן כללי כמה מושגים המגבילים את הביטוי הפנוטיפי של דמויות גנוטיפיות.
קודם כל נדבר על המושגים חדירה ואקספרסיביות, ואז נקדיש תשומת לב מיוחדת לתופעות של ויסות הפעולה הגנטית.
עמידות
חדירת הגן מייצגת את יכולתו להתבטא בפנוטיפ. חדירות נמדדת סטטיסטית על ידי ספירת תדירות הפנוטיפים המראים את האופי הזה מתוך 100 גנוטיפים המכילים אותו. תכונה עם 0.7 חדירות היא תכונה המתרחשת פנוטיפית ב -70% מהתדירות הגנוטיפית שלה.
ביטוי
אקספרסיביות היא הערכה כמותית של מידת הביטוי הפנוטיפי.
תקנת הפעולה הכללית
התאים מייצרים את כל האנזימים והחלבונים שלהם באותה מהירות ובאותו הזמן. למשל, ניתן לספק תאי Escherichia coli עם אנרגיה ואטומי פחמן מהדיסכריד הלקטוז מכיוון שהם מסוגלים לפרק אותם לגלוקוז וגלקטוז הודות לאנזים בטא-גלקטוסידאז. ב- E. coli רגיל שעלול להיות לקטוז, יש כ -3000 מולקולות של בטא-גלקטוסידאז, השוות ל -3% מהחלבונים של התא ההוא; בהעדר לקטוז תהיה רק מולקולה אחת של בטא-גלקטוסידאז לכל תא חיידקי. Galactosidase יסונתז ממולקולות mRNA חדשות כאשר ניתן יהיה להשתמש בו. זנים מוטנטים של E. coli העשירים באנזים ידועים גם כאשר נעדר לקטוז: מוטציות אלה נחלשות בהשוואה לתאים רגילים מכיוון שהם נאלצים לצרוך אנרגיה וחומרים מיותרים כדי לייצר את האנזים שיישאר ללא מצע. חומרים הגורמים לעלייה בכמות האנזים, כמו במקרה של הלקטוז, ייקראו אינדוזרים, בעוד שלאנזימים ניתן לומר שהם ניתנים להשראה. חומרים אחרים גורמים, גם אלה באופן ספציפי, לייצור אנזימים מסוימים. גם ב- E. coli, למשל, מסוגל לבנות את כל חומצות האמינו שלה, בעלות פחמן ואמוניום (NH3), נוכחות במדיום התרבות של חומצת אמינו מסוימת (היסטידין, למשל) חוסמת את ייצור כל האנזימים הקשורים ל- הביוסינתזה של חומצת האמינו עצמה: ייאמר על אנזימים אלה שהם ניתנים לדחיסה. בתאים חיידקיים נהרסים מולקולות ה- mRNA זמן קצר לאחר היווצרותם, ולכן שליטה על ייצור ה- mRNA פירושה שליטה בסינתזה האנזימטית באותו זמן זמן.
האופרון
להסביר כיצד התא החיידקי מסוגל לשלוט בייצור האנזימים שלו יעקב ומונוד גיבשו את השערת האופרון; האופרון נוצר על ידי מספר גנים הקשורים תפקודית ומיושרים ללא רצף לאורך קטע DNA. האופרון מורכב משלושה סוגים שונים של גנים: האמרגן, שם מתחיל היווצרות ה- mRNA; המפעיל, שבו מתבצעת שליטה; גן מבני אחד או יותר, המקודדים לאנזימים או לחלבונים אחרים. במערכת בטא-גלקטוסידאז, האופרון כולל, בנוסף לזה של בטא-גלקטוסידאז, גם שני גנים אחרים המקודדים מבנים עבור אנזימים אחרים המעורבים בחילוף החומרים של הלקטוז.גנים אלה צמודים זה לזה ומתועתלים אחד אחרי השני לאורך אותו סליל ה- DNA למולקולת mRNA אחת. מולקולות ה- mRNA שנוצרו כך פעילות לזמן קצר מאוד, ולאחר מכן הן נהרסות על ידי אנזימים ספציפיים..
הפעילות של האופרון נשלטת בתורו על ידי גן אחר, הרגולטור, שעשוי גם להיות רחוק מהאופרון: וסת זה מקודד חלבון, הנקרא רפרסור, אשר נראה כנקשר ל- DNA בגן המפעיל. בין האמרגן לגנים המבניים למעשה חוסם את ייצור ה- mRNA.
המדכא בתורו נשלט, והבקרה מתבצעת באמצעות חומר "אות". במקרה של אנזימים הניתנים להשראה חומר זה הוא "המשרה. המשרה" נקשר למולקולת הדכא המשנה את צורתו כך שלא יוכל להסתגל יותר לדנ"א: במקרה זה, מכיוון שאין רפרסור בין האמרגן לגנים המבניים. , הדכא יכול ליצור את מולקולות ה- mRNA ומתוכם את מולקולות החלבון. עם מיצוי שוב אספקת המשרן הרגולטור יחזיר לעצמו את השליטה, מה שיעצור את ייצור ה- mRNA החדש, ולכן של חלבונים חדשים. במערכת בטא-גלקטוסידאז המעורר הוא לקטוז או חומר הדומה מאוד לזה. נגזרת: הם יצטרף לרפרס שתפעיל אותו כך שיאפשר ביוסינתזה של אנזימים. במקרה של אנזימים הניתנים להדחקה, החומר שפועל כ"אות "פועל כמדכא: המדכא פעיל רק אם הוא משולב עם הלוח. במערכת ההיסטדין, הכוללת תריסר אנזימים שונים, זוהי חומצת אמינו זו, בשילוב עם ה- tRNA שלה, המחולל. Histidine.
אינטראקציות אלוסטריות
אינטראקציות אלוסטריות, הכרוכות בחוסר הפעלה של אנזים על ידי שינוי צורתו, מספקות דרך אחרת לווסת את הפעילות המטבולית של התא. אינטראקציות אלוסטריות מאפשרות שליטה מדויקת יותר ממערכת המשרן -מדכא של האופרון, אך אינן משיגות את התוצאה השימושית של אי הכללת הביוסינתזה של חומר נתון מהשלב הראשון - ייצור mRNA.
מערכות בקרה ב- EUCARIUS
יש כמה עובדות שמובילות להאמין שמערכת ויסות הדומה לאופרון פועלת ובולטת בקרב צמחים ובעלי חיים. הכרומוזומים של אורגניזמים אלה שונים באופן משמעותי מאלו של E. coli ופרוקריוטים אחרים. שליטת הגנים באלה התאים שונים מאוד.מנגנון המיטוזה הוא כזה שכל תא של צמח או בעל חיים נתון מחזיק בכל המידע
הגנטיקה קיימת בביצית המופרית. לכן רוב הגנים בכל תא מתמחה יישארו בלתי יעילים לאורך כל חיי התא. ה- DNA בתאים אלה קשור תמיד לחלבונים. לכן יתכן כי דיכוי גנים באיקריוטים מחייב בדיוק את הקשר הזה. בין DNA לחלבונים.