ההמשכיות המחזורית של יצורים חיים מוצאת את הקשרים בין דורות עוקבים בתופעות הרבייה.
הרבייה מתבצעת ברמות שונות של הסולם האבולוציוני, בהשלכות השונות של ממלכות הירקות ובעלי החיים, במינים החיים השונים, עם מגוון מנגנונים כל כך, אשר לבדם מצדיק מסה שלמה.
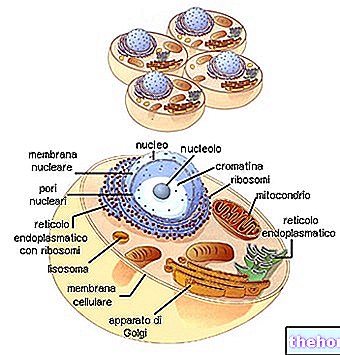
סיווג ראשון של תופעות הרבייה חייב להבחין בין חד -תאיים לבין אורגניזמים רב -תאיים, כיוון שרק באחרונה חלוקת התא עולה בקנה אחד עם הרבייה.
ברב תאיים, רבייה יכולה להיות אגמית או מינית (או גמית).
רבייה אגמית, שכיחה יחסית, מבוססת על מנגנון המיטוזה, כך שהשונות של המין מופקדת דווקא על התרחשות חוזרת של מוטציות.
ישנם גם מנגנונים שונים, כגון strobilization, sporulation, וכו ', בעוד בצמחים אנו מוצאים צורות התחדשות מוכרות היטב בחקלאות (ייחורים, שכבות וכו').
אולם מנגנון הרבייה הנפוץ ביותר בצורות הגבוהות יותר הוא המין, המתאים להופעת מיוזה, להיווצרות גמטות ולהתמזגותן בזיגוטה (הפריה).
במינים פרימיטיביים הגמטות אינן מובדנות מבחינה מורפולוגית: במקרה זה אנו מדברים על איזוגמיה. עם זאת, תמיד ישנן שתי סדרות של גמטות, המזוהות עם הסמלים (+) ו- (-) וההפריה יכולה להתרחש רק מתוך "המפגש בין גמטות של סימן הפוך: לכן יש הבדל ביולוגי, שטרם בא לידי ביטוי מבחינה מורפולוגית.
ככל שהסולם האבולוציוני מתקדם, מופיעה התמיינות מורפולוגית ופונקציונלית, שבה סוג של גמטות נקבות ניחן בדרך כלל בחומר עתודה בשפע (הדאוטופלזמה או העגל, שיבטיחו את התפתחות העובר עד שיהיה עצמאי מבחינה מטבולית) וסוג של גמטות זכריות ניחנו בתנועתיות להגיע לנשים. גמטות הן תמיד הפלואידיות והן תוצאה של מיוזה. ההתמזגות שלהן גורמת לזיגוטה הדיפלואידית.
בין מיוזיס לזיגוטה יכולים לחלוף סדרת דורות של תאים הפלואידים, כשם שבין זיגוטה למיוזה יכולים לחלוף סדרת דורות של תאים דיפלואידים, כאשר הזנים השונים של מחזורי יצירה מתבטאים בחילופי הדורות.
מין דיפלונט (בעל אורגניזם דיפלואידי) מאופיין במיוזה גמטית: מיוזה מייצרת ישירות את הגמטות, אשר על ידי מיזוג מייצרות מחדש את המצב הדיפלואידי באופן מיידי. זהו המקרה הרווח של המטאואנים, כולל האדם.
גמטוגנזה
בהתחשב ברבייה עם מיוזה גמטית, כפי שהיא מתרחשת באדם, בואו ננסה להבהיר כיצד מיוזה משתלבת בגמטוגנזה (היווצרות גמטות).
בהתפתחות העובריים, של גמטוגנזה זכר ונקבה (הנקראים זרעי זרע ואוגנזה), קיימת התמיינות מוקדמת בין התאים שנועדו ליצור את הגוף (קו סומטי) לבין אלה המיועדים לייצר גמטות (קו נבט). התאים הראשוניים של קו הנבט נקראים פרוטוגונים. עם התמיינות של בלוטת המין במובן הגברי או הנקבי ישנה התמיינות של תאי הנבט בהתאמה לספרמטוגוניה ואובוגוניה.
במבט על הזרע, אנו רואים שיש סדרה של דורות תאים בספרמטוגוניה, הנמשכת לאורך כל החיים. רק חלק מהפרמטוגוניה המיוצרת באופן רציף שונה מהמחזור המיטוטי הרגיל והמיוטי מתחיל במקום זאת.
תא הנבט בו תתחיל מיוזה (צמצום ולאחר מכן חלוקה ראשונה) נקרא זרע ראשון (spermatocyte); חלוקתה מולידה שני זרעי זרע מסדר שני, אשר עם החלוקה השנייה מולידים סך של ארבעה זרעונים.
אנו יכולים לבחון את ההפחתה של ערכת הכרומוזומלים מ 4n של הזרע הראשון (לאחר צמצום ישנם ארבעה כרומטילים לכל זוג הומולוגים) ל- 2n של הזרע השני של הזרע ול- n של הזרעונים, כפי שכבר נראה על ידי מחקר מיוזה, אשר כך אנו מסיקים. הזרעים הם אפוא כבר הפלואידים, אך הם עדיין לא גמטות בוגרות. ממבנה של תא מסוג הפלואידי, התבגרות תפקודית (הנקראת spermiohistogenesis) הופכת את הזרעונים לזרעונים בוגרים.
בגמטוגנזה נשית (או אווגנזה) ישנם מספר הבדלים. קודם כל, מספר הגאמטות שיש להכין קטן בהרבה. ההערכה היא שכ -5 X 105 אובוגונים מוכנים בגונדות של נקבה ממין האדם; מתוכם, רק כ -400 מתעניינים בהבשלה של זקיקים ובדכאון שלאחר מכן, במחזור שלרוב משפיע על זקיק אחד בלבד בחודש לתקופה פורייה של כ -35 שנים.
מספר הגאמטות השונות שהוכנו בשני המינים תואם את ההבדל בתפקוד ובהתנהגות שכבר הוזכרו: הזרעונים קטנים, ניידים ורבים ביחס לצורך בחיפוש הביצה וההסתברות הנמוכה למצוא אותה; הביצים הן גדול, אינרטי ומעט, ביחס לתפקוד של הבטחת העובר לחומר המילואים וההגנה הניתנת להם על ידי הפריה פנימית (באופן טבעי, במיוחד בהפריה חיצונית גם הביציות חייבות להיות רבות יותר).
הצורך לספק לגאמטים חומר עתודה תואם לנוכחות, ב"אוגנזה ", שלב של עצירת מיוזה, שבמהלכו הסרטן חלקי של הכרומוזומים. לאחר מכן אנו מתבוננים במה שנקרא" כרומוזומי נוצה ", בהם סדרה של מוחצנים מזהה את התכונות בהן הדיכאון של הגנים האחראים לסינתזה של דיאוטופלזמה.
המספר הקטן יותר הנדרש לגמטות נקבות תואם גם את העובדה שמבין ארבעת התאים הפלואידים המיוצרים על ידי מיוזה, רק אחד מקבל את כל חומר המילואים והופך לגאמט, ואילו שלושת האחרים (הפולוציטים או גופי הקוטב), המכילים רק חומר כרומוזומלי, אינו יכול להוליד זיגוטים ועוברים והם מיועדים לסגת.
הַפרָיָה
הפריה, כלומר המפגש בין גמטה זכר לנקבה, יכולה להתבצע בדרכים שונות מאוד.בממלכת החיות אנו רואים את המעבר מהפרייה חיצונית (גמטות שנחשפות לכל סיכון סביבתי ולכן בהכרח רבות מאוד בשני המינים) להפריה פנימית, אליה קשורה טיפול ההורים עוד יותר ליחסים המטבוליים של היונקים בין האם לעובר. .
הפריה, לאחר שהמפגש בין גמטות מהמין השני מתרחש, חייב להתקיים המבטיח שני תנאים: ייחודיות וייחודיות. כלומר, יש לוודא שהזרע הוא מאותו מין כמו הביצה ושברגע שהראשון נכנסת, אף אחד לא נכנס.
הספציפיות מובטחת על ידי המאפיינים הביוכימיים של האקרוזום ומשטח הביצית. למעשה, יש דיבורים על תגובות בין «פורטיליזינים» ל- «אנטי -פוריות», כאשר הספציפיות שלהן דומה לזו של המפגש בין אנזימים ומצע.
ייחודיות ההפריה מובטחת על ידי שינוי מבנה פני השטח של הביצית ("תגובה קליפת המוח") המתחיל ממש ברגע התגובה הספציפית הראשונה של הפרוטיליסין / אנטי -פרטיצין; לאחר תגובה זו, קרום הביצית משתנה, כך שכל זרע אחר שיגיע אליו כבר לא יוכל ליזום את תגובת ההפריה הספציפית.
לאחר ההפריה, זנבו של הזרע נותר חיצוני לביצית, בעוד שהחומר הכרומוזומלי חודר לתוכו. זה, המכונה "פרונוקלאוס זכר", מצטרף ל"פרונוקלאוס הנשי "של הביצית ובכך יוצר את הגרעין הדיפלואידי של הזיגוטה.