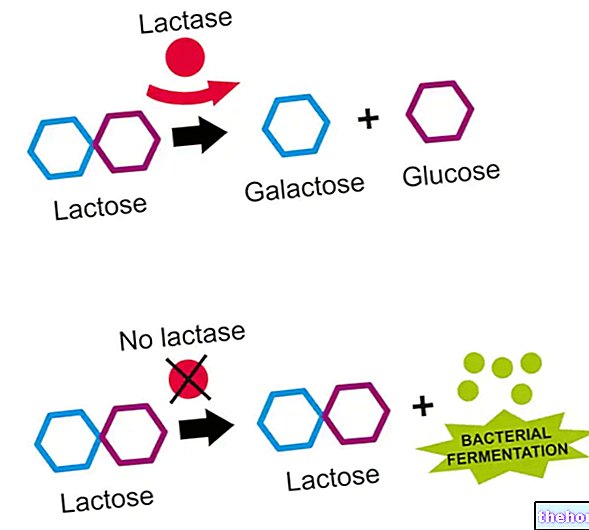
אם תא פועל בתנאים אנאירוביים, הוא מייצר אנרגיה על ידי הפיכת גלוקוז ללקטט, ובאמצעות מחזור הקורי, הוא זורק את זה; אם יש חמצן זמין (ולכן בתנאי מנוחה), יותר מ -90% מהגלוקוז נצרך באופן אירובי ו רק 10%הנותרים, אנאירובית. כאשר יש צורך ביותר ATP ממה שהמסלול האירובי מסוגל לספק (למשל כאשר השרירים נמצאים במתח), אז האספקה הנוספת מסופקת על ידי מטבוליזם אנאירובי (אנו נמצאים במצבי מחסור בחמצן: קוצר נשימה, עייפות בנשימה וכו '): יש צורך להאיץ את חילוף החומרים על ידי הפיכת לקטט (המתקבל מגליקוליזה) לגלוקוז באמצעות גלוקונאוגנזה.
חילוף החומרים האירובי מתפתח במיטוכונדריה.
האנזים הראשון שנתקל בחילוף החומרים האירובי הוא פירובט דהידרוגנאז; יותר נכון לומר שפירובאט דהידרוגנאז הוא קומפלקס אנזים ולא אנזים, מכיוון שהוא אגרגט של 48-60 יחידות חלבון עם שלושה אתרים קטליטיים הפועלים ברצף.
פירובט דהידרוגנאז מזרז את התגובה הבאה (חיזור):
פירובט + NAD + + CoA-SH → אצטיל CoA + NADH + H + + CO2
CoA-SH הוא קואנזים A: הוא נגזרת של חומצה פנטותנית; אצטיל קואנזים A הוא תיאוסטר. זהו תהליך חמצון מכיוון שהפחמן הראשון של פירובאט עובר מחמצון מספר שלוש לחמצון מספר ארבע (הוא מתחמצן) והפחמן השני של פירובאט עובר מחמצון מספר שתיים לחמצון מספר שלוש (הוא מתחמצן). ואז הפירובאט מתחמצן (הוא מאבד שני אלקטרונים בסך הכל) וה- NAD מצטמצם.
כאמור, לפירובאט דהידרוגנאז יש שלושה סוגים של פעילות אנזימטית, כל אחד נתמך על ידי קופקטור קטליטי משלו:
- תיאמין פירופוספט (הוא נגזרת של ויטמין B1); הוא פעיל בצורה מופחתת: קרבוניון נוצר.
- ליפואמיד (הוא נגזרת של חומצה ליפואית); הוא מכיל גשר דיסולפידי תגובתי מאוד.
- flavin adenin dinucleotide (זהו נגזרת של ויטמין B2); זהו נוקלאוטיד בעל תכונות חמצון: מרכז החיזור שלו מורכב מהפלבין.
בתאים אוקריוטים, מטבוליזם אירובי מתרחש באברונים מיוחדים של התא שהם המיטוכונדריה; בחיידקים חילוף החומרים של גלוקוז ומינים אחרים מתרחש בתא אך אין אברונים מיוחדים.
כאשר פירובאט נכנס למיטוכונדריון, הוא נתון ל"פעולה של פירובקט קרבוקסילאז אם יש צורך לבצע גלוקונאוגנזה (לשחזור החומר ההתחלתי), או שהוא יכול להיות נתון לפירובאט דהידרוגנאז אם יש צורך לייצר אנרגיה: "אצטיל קואנזים A הנוצר על ידי מטבוליזם אירובי מגרה את פעולתו של פירובט קרבוקסילאז, לכן הוא מקדם את הגלוקוקונאוגנזה ומפחית את פעולתו של פירובט דהידרוגנאז.
בואו נראה כעת כיצד פועל פירובט דהידרוגנאז; קודם כל, יש decarboxylation של pyruvate על ידי הפעולה של pyrophosphate תיאמין.
סביבה חומצית יכולה לעכב מטבוליזם אירובי מכיוון שהצורה האוניונית של תיאורון פירופוספט פעילה אשר תועתק ב- pH חומצי ו decarboxylation לא תתרחש.
דקרבוקסילציה היא תגובה קשה מכיוון שצריך לשבור קשר פחמן-פחמן; במקרה זה התגובה מועדפת מבחינה תרמודינמית על ידי העובדה שתגובת התגובה (hydroxyethyl-thiamine pyrophosphate) מעניקה תהודה (האלקטרונים p של המולקולה מופקים): הפירופוספט הידרוקסיאתיל-תיאמין קיים בשלוש צורות אפשריות (תהודה) ו- זה הופך אותו ליציב למדי. יתר על כן, הפירופוספט הידרוקסיאתיל-תיאמין בצורת אניון שורד מספיק זמן מספיק כדי להיות מסוגל לקיים אינטראקציה עם גשר הדיסולפיד של הליפואמיד (הקופקטור הקטליטי השני של פירובט דהידרוגנאז); גשר הדיסולפיד הוא זרוע מתנדנדת (הוא ממוקם על סוף שרשרת גמישה וארוכה) ויכול לעבור מאתר קטליטי אחד לשני במתחם האנזים.
ואז הליפואמיד, דרך גשר הדיסולפיד, קושר את הפירופוספט הידרוקסיאתיל-תיאמין: מתקבל אצטיל ליפואמיד.זה שתואר זה עתה הוא השלב הראשון בתגובת transacetylation המזרזת על ידי האנזים הראשון של מכלול pyruvate dehydrogenase; בשלב זה הופר קשר בין קבוצת ההידרוקסיל לפירופוספט תיאמין שחזר לצורתו המקורית: התקיימה תגובת חמצון שבה גשר הדיסולפיד פעל כחמצון (שני אטומי הגופרית מופחתים) בהשוואות בקבוצת ההידרוקסיל אשר מחומצן לאצטיל.
לאחר שלב זה, הזרוע המתנדנדת של הליפואמיד נעה ומתקרבת לאנזים השני של פירובט דהידרוגנאז המבצע את פעילות הטרנסאצטילאז האמיתית על ידי נשיאת עמו קבוצת האצטיל: מתקיים השלב השני של תגובת הטרנסצילציה המזרזת על ידי האנזים השני; בדרך זו השגנו אצטיל קואנזים א. כעת יש צורך לשחזר את הליפואמיד הנמצא בצורה מופחתת: האנזים השלישי של פירובאט דהידרוגנאז מתערב אשר מחמצן מחדש את הליפואמיד ומעביר את האלקטרודות שלו ל- FAD אשר מצטמצם ל- FADH2. בני הזוג. FAD / FADH2 יכול לתפקד כזוג חמצון בשני שלבים חד -אלקטרוניים מובחנים או בשלב דו -אלקטרוני יחיד.
ה- FADH2 נותן מיד את האלקטרונים שלו ל- NAD + ומקבל FAD ו- NADH + H +.
אצטיל קואנזים A, המתקבל כמתואר, הוא המוצר ההתחלתי למחזור קרבס (או מחזור של חומצות טריקרבוקסיליות).


.jpg)