נערך על ידי ד"ר ג'ובאני צ'טה
מביוכימיה ועד ביומכניקה

מטריצה חוץ-סלולרית (MEC)
תיאור, אף על פי המעט שאנו מכירים כיום, של ה- ECM (מטריצה חוץ -תאית) חיוני כדי להבין טוב יותר את חשיבות השינויים בעמוד השדרה והיציבה בבריאות.
כל תא, כמו כל אורגניזם חי רב תאיים, צריך "להרגיש" ולתקשר עם סביבתו על מנת לבצע את תפקידיו החיוניים ולשרוד. באורגניזם רב תאי התאים חייבים לתאם את ההתנהגויות השונות כמו בקהילה של בני אדם.

ECM מתואר בדרך כלל כמורכב מכמה סוגים גדולים של ביו -מולקולות:
- חלבונים מבניים (קולגן ואלסטין)
- חלבונים מיוחדים (פיברילין, פיברונקטין, למינין וכו ')
- פרוטוגליקנים (אגרקנים, סינדקנים) וגלוסאמינוגליקנים (היאלורוננים, כונדרואיטין סולפטים, גופריות הפארן וכו ')
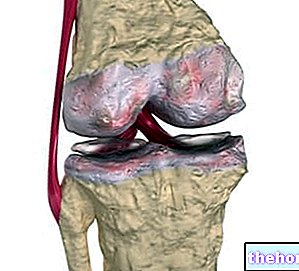
בין החלבונים המבניים, כלומר קולגן יוצרים את משפחת הגליקופרוטאינים המיוצגת ביותר בממלכת החיות. הם החלבונים הקיימים ביותר ב- ECM (אך לא החשובים ביותר) והם המרכיבים הבסיסיים של רקמות החיבור הנכונות (סחוס, עצם, פאשיה, גידים, רצועות).
קולגנים מסונתזים ברובם על ידי פיברובלסטים אך תאי אפיתל מסוגלים גם לסנתז אותם.
סיבי הקולגן פועלים באופן רציף עם כמות עצומה של מולקולות אחרות של ה- ECM המהוות רצף ביולוגי מהותי לחיי התא. הקולגנים הקשורים בסיבים תופסים תפקיד דומיננטי ביצירה ותחזוקה של מבנים המסוגלים להתנגד לכוחות המתח כמעט קולגן לא אלסטי מיוצר ומטבוליזם מחדש כפונקציה של העומס המכני ולתכונותיו הוויסקו-אלסטיות יש השפעה רבה על יציבת האדם.
סיבי הקולגן הודות לציפוי ה- PG / GAG שלהם (פרוטאוגליקנים / גלוקוזאמינוליקנים) הם בעלי תכונות ביו -חיישניות ומוליכות ביולוגיות. אנו יודעים למעשה שכל כוח מכני המסוגל ליצור עיוות מבני מדגיש את הקשרים הבין-מולקולריים המייצרים שטף חשמלי קל, כלומר הזרם הפיזואלקטרי (Athenstaedt, 1969). לכן גם לרשת הקולגן התלת מימדית ובכל מקום יש את המאפיין המיוחד של העברת אותות ביואלקטריים בשלושת ממדי החלל, המבוססים על הסידור היחסי בין סיבי קולגן לתאים, בכיוון האפירנטי (מה- ECM לתאים) או להפך להפך.
כל זה מייצג מערכת תקשורת תאי MEC בזמן אמת ואותות ביו אלקטרומגנטיים כאלה יכולים להוביל לשינויים ביוכימיים חשובים, למשל, ב"אוסטאוקלאסטים בעצמות אינם יכולים "לעכל" עצם טעונה פיזואלקטרית (אושמן, 2000).
לבסוף, יש להדגיש כי התא, באופן לא מפתיע, מייצר באופן רציף ועם הוצאה ניכרת של אנרגיה (כ -70%) חומר שיש בהכרח לגרש באמצעות אחסון בלעדי של פרוטוקולגן (מבשר ביולוגי של קולגן) בשלפוחיות ספציפיות ( אלברגטי, 2004).
הרוב המכריע של רקמות החוליות דורשות נוכחות בו זמנית של שני מאפיינים חיוניים: חוזק וגמישות. רשת אמיתית של סיבים אלסטיים, הממוקם בתוך ה- ECM של רקמות אלה, מאפשר לחזור לתנאים הראשוניים לאחר עיכובים חזקים. הסיבים האלסטיים מסוגלים להגדיל את התרחבות האיבר או חלק ממנו לפחות חמש פעמים. סיבי קולגן ארוכים ובלתי אלסטיים נחלבים בין הסיבים האלסטיים במטרה מדויקת להגביל "עיוות יתר על ידי משיכה של הרקמות." אלסטין מייצג את המרכיב העיקרי של סיבים אלסטיים ונמצא בכמויות גדולות במיוחד בכלי דם בעלי מאפיינים אלסטיים ( מהווה יותר מ -50% מהמשקל היבש הכולל של אבי העורקים), ברצועות, בריאה ובעור.
ECM מכיל מספר גדול (ועדיין לא מוגדר היטב) של חלבונים מיוחדים שאינם קולגן, בעלי בדרך כלל אתרי קישור ספציפיים למולקולות ECM אחרות וקולטני משטח התא. בדרך זו, כל מרכיב בודד של החלבונים הללו פועל כ"מגבר "של קשרים, בין מולקולות דומות ושונות, ויוצר רשת ביוכימית אינסופית המסוגלת לייצר, לאפנן, להשתנות ולהתפשט גם למרחקים של מיליוני ומיליוני מידע ביוכימי. (ואנרגיה).
חלבון "מיוחד" של המטריצה החוץ -תאית הוא פיברונקטין, גליקופרוטאין בעל משקל מולקולרי גבוה המצוי בכל החוליות. נראה כי פיברונקטין מסוגל להשפיע על צמיחת תאים, הדבקה בין תאית ועם ECM, נדידת תאים בדרכים שונות (התא יכול לנוע עד 5 ס"מ ביום - אלברגטי, 2004) וכו 'האיזופורם הידוע ביותר, סוג III, נקשר לאינטגרינים . האחרונים הם משפחה של חלבונים טרנסממברניים הפועלים כקולטנים מכניים: הם מעבירים, באופן סלקטיבי ובאופן הניתן לשינוי, פעולות מכניות ודחיפות מה- ECM בתוך התא ולהיפך, וגורמות לסדרה של תגובות בציטופלזמה הכוללות את השלד וה חלבונים אחרים שהם מווסתים את הידבקות התא, הצמיחה וההגירה (Hynes, 2002).
Glucosaminoglycans (GAGS) ו- proteoglycans (PGs) יוצרים חומר דמוי ג'ל מיובש מאוד המוגדר בתוך רקמות החיבור, שבתוכו נמצאים חלבוני הפיברילרים ושומרים עליהם. צורה זו של ג'ל פוליסכריד מסוגלת מצד אחד לאפשר ל- ECM לעמוד בפני כוחות דחיסה ניכרים ומצד שני לאפשר דיפוזיה מהירה, קבועה וסלקטיבית של חומרים מזינים, מטבוליטים והורמונים בין דם לרקמות.
שרשראות הפוליסכרידים של הגלוקוזמינוגליקנים נוקשות מכדי לקפל בתוך המבנים הכדוריים הקומפקטיים האופייניים לשרשראות פוליפפטיד, בנוסף הם הידרופיליים ביותר. מסיבות אלה (וכנראה גם עבור אחרים שאינם ידועים לנו) ה- GAG נוטים להניח קונפורמציות קיצוניות ביותר. תופסים נפח גדול ביחס למסתם ובכך יוצרים כמויות ניכרות של ג'ל גם בריכוזים נמוכים. הכמות הגבוהה של מטענים שליליים (ה- GAGs מייצגים את התאים האניוניים הרבים ביותר, בדרך כלל מנופחים, המיוצרים על ידי תאי בעלי חיים) מושכת קטיונים רבים; בין אלה תפקיד דומיננטי ממלא Na + שנותן את היכולת האוסמוטית כולה ולוכד כמות עצומה של מים ב- ECM. בדרך זו נוצרים נפיחות (טורגורים) המאפשרים ל- ECM להתנגד אפילו לכוחות דחיסה חשובים (הודות לכך, למשל, סחוס הירך יכול, בתנאים פיזיולוגיים, להתנגד בצורה מושלמת ללחץ של כמה מאות אטמוספרות).
בתוך רקמת החיבור, ה- GAGs מייצגים פחות מ-10-12% מהמשקל הגלובלי, אולם הודות למאפייניהם הם ממלאים רבים מהחללים החוץ-תאים ויוצרים נקבוביות של ג'ל לחות בגדלים וצפיפויות שונות של מטענים חשמליים ובכך פועלים כסלקטיביים. נקודות מפתח או "שרתים" שדרכם מוסדרת תנועת המולקולות והתאים בתוך ה- MEC, בהתבסס על גודלם, משקלם ומטען החשמלי.
חומצה היאלורונית (היאלורונאן, היאלורונאט) מייצגת אולי הפשוטה מבין ה- GAG. נתונים ניסיוניים ומולקולריים-ביולוגיים מאשרים כי היא ממלאת תפקיד מהותי ברמת העצמות והמפרקים בנוגע לעמידות בפני לחצים ניכרים. מילוי חללים ב- ECM במהלך התפתחות עוברית : הוא יוצר רווחים ריקים בין התאים שאליהם הם ינועו בשלבים מאוחרים יותר (Albergati, 2004).
לא כל ה- PG מופרשים על ידי ECM, חלקם הם מרכיבים אינטגרליים של ממברנות הפלזמה (Alberts, 2002).
מטריקס חוץ-סלולרי יכול אפוא להיחשב כרשת מורכבת מאוד שבה חלבונים, PGS ו- GAG מספקים אינספור פונקציות כולל תמיכה מבנית וויסות כל רקמה ופעילות אורגנית. יש להתייחס להומאוסטזיס סלולרי גלובלי כמכלול של מנגנונים שיכולים להתפתח ולהתפתח בתוך התא או מבחוץ ב- ECM; במקרה האחרון התא יכול לייצג את המטרה הבינונית או הסופית. המרכיבים החוץ -תאים, בנוסף לייצוג מבני תמיכה פיזיים לפיגום התאי, משמשים גם כאתרים אמיתיים ליזום, פיתוח וסיום של תהליכים חיוניים הנוגעים הן לסביבה האנדו -תאית והן לאיברים ולמערכות. אנו ניצבים בפני רשת ביוכימית אינסופית המסוגלת לייצר, לאפנן, לשנות ולהפיץ, אפילו מרחוק, מיליוני ומיליוני מידע.
כל תא בגוף מתקיים כל הזמן עם ECM, הן בהיבטים המכניים והכימיים והן האנרגטיים, עם השפעות "דרמטיות" על הארכיטקטורה הסטטית והדינאמית של הרקמות. לדברי פ.א.א באצ'י, המטריצה הביניים מייצגת באמת את אם התגובות החיוניות, המקום שבו, קודם כל, מתקיימים חילופי הדברים בין החומר לאנרגיה. כל הרקמות מחוברות ומשולבות זו בזו מבחינה תפקודית לא במערכות סגורות אך פתוחות; מתקיימים ביניהם חילופים רציפים, שיכולים להתרחש הן מקומית והן מערכתית, תוך ניצול מסרים ביוכימיים, ביופיזיאליים ואלקטרומגנטיים, כלומר שימוש בצורות האנרגיה השונות.
כפי שאומר פ.ג. אלברגטי, התא והמטריצה החוץ -תאית מייצגים שני עולמות נפרדים לכאורה שבהכרח לכל משך החיים חייבים לקיים אינטראקציה בכל רגע כדי לפעול בצורה נכונה וסינרגטית. זה דורש סדרת אותות יוצאת דופן ואחריה סדרה לא פחות מדהימה של פעילויות מולקולריות-ביולוגיות.
מאמרים נוספים בנושא "מטריצה חוץ -סלולרית - מבנה ופונקציות"
- טיפול בעקמת
- עקמת - סיבות ותוצאות
- אבחון עקמת
- פרוגנוזה של עקמת
- רקמת חיבור ופשיה חיבורית
- רצועת חיבור - תכונות ופונקציות
- יציבה ומתיחות
- תנועת האדם והחשיבות של תמיכת עכוז
- החשיבות של עכוז ותומכים נכונים
- עקמת אידיופטית - מיתוסים להפיג
- מקרה קליני של עקמת ופרוטוקול טיפולי
- תוצאות הטיפול עקמת קלינית
- עקמת כגישה טבעית - ביבליוגרפיה